Medicinski strokovnjak članka

Nove publikacije
Jajčnik
Zadnji pregled: 04.07.2025

Vsa vsebina iLive je pregledana ali preverjena, da se zagotovi čim večja dejanska natančnost.
Imamo stroge smernice za pridobivanje virov in samo povezave do uglednih medijskih strani, akademskih raziskovalnih institucij in, kadar je to mogoče, medicinsko pregledanih študij. Upoštevajte, da so številke v oklepajih ([1], [2] itd.) Povezave, ki jih je mogoče klikniti na te študije.
Če menite, da je katera koli naša vsebina netočna, zastarela ali drugače vprašljiva, jo izberite in pritisnite Ctrl + Enter.
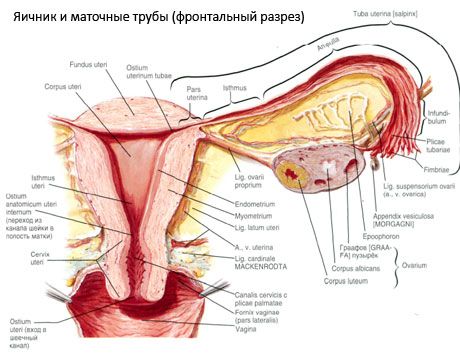
Jajčnik (ovarium; grško oophoron) je parni organ, ženska spolna žleza, ki se nahaja v medenični votlini za širokim ligamentom maternice. V jajčnikih se razvijajo in dozorevajo ženske spolne celice (jajčeca), nastajajo pa tudi ženski spolni hormoni, ki vstopajo v kri in limfo. Jajčnik ima jajčasto obliko, nekoliko sploščen v anteroposteriorni smeri. Barva jajčnika je rožnata. Na površini jajčnika ženske, ki je rodila, so vidne vdolbine in brazgotine - sledi ovulacije in preobrazbe rumenih telesc. Jajčnik tehta 5-8 g. Dimenzije jajčnika so: dolžina 2,5-5,5 cm, širina 1,5-3,0 cm, debelina - do 2 cm. Jajčnik ima dve prosti površini: medialno površino (facies medialis), obrnjeno proti medenični votlini, delno prekrito z jajcevodom, in stransko površino (facies lateralis), ki meji na stransko steno medenice, do rahlo izražene vdolbine - jajčnikovo jajčnično jamico. Ta fosa se nahaja v kotu med zunanjimi iliakalnimi žilami, ki so na vrhu prekrite s peritoneumom, in materničnimi in obturatornimi arterijami na dnu. Za jajčnikom retroperitonealno od zgoraj navzdol poteka ureter ustrezne strani.
Površine jajčnika prehajajo v konveksen prosti (zadnji) rob (margo liber), spredaj pa v mezenterični rob (margo mesovaricus), ki je s kratko gubo peritoneuma (mezenterija jajčnika) pritrjen na zadnji listič širokega ligamenta maternice. Na tem sprednjem robu organa je žlebasta vdolbina - hilum jajčnika (hilum ovarii), skozi katerega v jajčnik vstopajo arterija in živci, iz njega pa izstopajo vene in limfne žile. Jajčnik ima tudi dva konca: zaobljen zgornji cevasti konec (extremitas tubaria), obrnjen proti jajcevodu, in spodnji maternični konec (extremitas utenna), ki ga z maternico povezuje jajčnikov ligament (lig. ovarii proprium). Ta ligament v obliki okrogle vrvice, debele približno 6 mm, poteka od materničnega konca jajčnika do stranskega kota maternice, ki se nahaja med obema listoma širokega ligamenta maternice. Ligamentozni aparat jajčnika vključuje tudi ligamentni suspenzor jajčnika (lig.suspensorium ovarii), ki je guba peritoneuma, ki poteka od stene male medenice do jajčnika in vsebuje jajčne žile ter snope vlaknatih vlaken. Jajčnik je pritrjen s kratkim mezenterijem (mesovarium), ki je podvojitev peritoneuma in poteka od zadnjega lističa širokega ligamenta maternice do mezenteričnega roba jajčnika. Sami jajčniki niso pokriti s peritoneumom. Največji jajčni rob jajcevoda je pritrjen na cevasti konec jajčnika. Topografija jajčnika je odvisna od položaja maternice in njene velikosti (med nosečnostjo). Jajčniki so zelo gibljivi organi male medenične votline.
 [
[ Plovila in živci jajčnikov
Oskrbo jajčnikov s krvjo zagotavljajo aa. et vv. ovaricae et uterinae. Obe jajčnikovi arteriji (aa. ovaricae dextra et sinistra) izvirata na sprednji površini aorte tik pod ledvičnimi arterijami; desna običajno izvira iz aorte, leva pa iz ledvične arterije. Vsaka jajčnikova arterija, ki poteka navzdol in lateralno vzdolž sprednje površine velike ledvične mišice, prečka sečevod spredaj (od njega oddaja veje), zunanje iliakalne žile, mejno črto in vstopi v medenično votlino, kjer se nahaja v suspenzornem ligamentu jajčnika. V medialni smeri jajčnikova arterija prehaja med listi širokega ligamenta maternice pod jajcevodom, od njega oddaja veje, nato pa v mezenterij jajčnika; vstopi v hilum jajčnika.
Veje jajčne arterije se široko anastomozirajo z jajčnimi vejami maternične arterije. Venski odtok iz jajčnikov poteka predvsem v jajčni venski pleksus, ki se nahaja v predelu jajčnega hiluma. Od tod odtok krvi poteka v dveh smereh: skozi maternične in jajčne vene. Desna jajčna vena ima zaklopke in se izliva v spodnjo votlo veno. Leva jajčna vena se izliva v levo ledvično veno, čeprav nima zaklopk.
Limfna drenaža iz jajčnikov poteka po limfnih žilah, še posebej obilno na območju organskih vrat, kjer se razlikuje subovarijski limfni pleksus. Nato se limfa po limfnih žilah jajčnikov odvaja v paraaortalne bezgavke.
Inervacija jajčnikov
Simpatični - zagotavljajo ga postganglijska vlakna iz celiakalnega (sončnega), superiornega mezenteričnega in hipogastričnega pleksusa; parasimpatični - visceralni sakralni živci.
Struktura jajčnika
Površina jajčnika je prekrita z enoslojnim zarodnim epitelijem. Pod njo leži gosta vezivno-tkivna beljakovinska ovojnica (tunica albuginea). Vezivno tkivo jajčnika tvori njegovo stromo (stroma ovarii), bogato z elastičnimi vlakni. Snov jajčnika, njegov parenhim, je razdeljena na zunanjo in notranjo plast. Notranja plast, ki se nahaja v središču jajčnika, bližje njegovim vratom, se imenuje medula (medulla ovarii). V tej plasti, v rahlem vezivnem tkivu, so številne krvne in limfne žile ter živci. Zunanja plast jajčnika - skorja (cortex ovarii) je gostejša. Vsebuje veliko vezivnega tkiva, v katerem se nahajajo dozorevajoči primarni jajčni folikli (folliculi ovarici primarii), sekundarni (vezikularni) folikli (folliculi ovarici secundarii, s.vesiculosi), pa tudi zreli folikli, Graafovi folikli (folliculi ovarici maturis), pa tudi rumena in atretična telesca.
Vsak folikel vsebuje žensko reproduktivno jajčece ali oocit (ovocytus). Jajčece ima premer do 150 µm, je okroglo, vsebuje jedro, veliko količino citoplazme, ki poleg celičnih organelov vsebuje tudi beljakovinsko-lipidne vključke (rumenjak) in glikogen, ki je potreben za prehrano jajčeca. Jajčece običajno porabi svojo zalogo hranil v 12–24 urah po ovulaciji. Če do oploditve ne pride, jajčece umre.
Človeško jajčece pokrivata dve membrani. V notranjosti je citolema, ki je citoplazemska membrana jajčeca. Zunaj citoleme je plast tako imenovanih folikularnih celic, ki ščitijo jajčece in opravljajo funkcijo tvorbe hormonov – izločajo estrogene.
Fiziološki položaj maternice, jajcevodov in jajčnikov zagotavljajo viseči, fiksirni in podporni aparati, ki združujejo peritoneum, vezi in tkivo medenice. Viseči aparat predstavljajo parne tvorbe, ki vključujejo okrogle in široke vezi maternice, lastne vezi in viseče vezi jajčnikov. Široke vezi maternice, lastne in viseče vezi jajčnikov držijo maternico v srednjem položaju. Okrogle vezi vlečejo fundus maternice naprej in zagotavljajo njegov fiziološki nagib.
Fiksirni (sidrni) aparat zagotavlja položaj maternice v središču majhne medenice in praktično onemogoča njeno premikanje vstran, naprej in nazaj. Ker pa se ligamentni aparat v spodnjem delu maternice oddaljuje od maternice, so možni nagibi maternice v različne smeri. Fiksirni aparat vključuje vezi, ki se nahajajo v ohlapnem tkivu medenice in segajo od spodnjega dela maternice do stranske, sprednje in zadnje stene medenice: sakromagične, kardinalne, uterovezikalne in vezikopubične vezi.
Poleg mezovarija se razlikujejo še naslednji ligamenti jajčnikov:
- suspenzorni ligament jajčnika, prej imenovan infundibulopelvični ligament. Gre za gubo peritoneuma s krvnimi žilami (a. et v. ovarica) ter limfnimi žilami in živci jajčnika, ki potekajo skozenj, raztegnjeno med stransko steno medenice, ledveno fascijo (na območju delitve skupne iliakalne arterije na zunanjo in notranjo) in zgornjim (tubalnim) koncem jajčnika;
- Prava vez jajčnika poteka med listi široke maternične vezi, bližje zadnjemu listu, in povezuje spodnji konec jajčnika s stranskim robom maternice. Prava vez jajčnika je pritrjena na maternico med začetkom jajcevoda in okroglo vezjo, za in nad slednjo. V debelini vezi potekajo rr. ovarii, ki so končne veje maternične arterije;
- Apendikularno-jajčni ligament Ligament se razteza od mezenterija slepiča do desnega jajčnika ali širokega ligamenta maternice v obliki peritonealne gube. Ligament je nestalen in ga opazimo pri 1/2 - 1/3 žensk.
Podporni aparat predstavljajo mišice in fascija medeničnega dna, razdeljene na spodnjo, srednjo in zgornjo (notranjo) plast.
Najmočnejša je zgornja (notranja) mišična plast, ki jo predstavlja parna mišica, ki dviguje anus. Sestavljena je iz mišičnih snopov, ki se pahljačasto raztezajo od trtice do medeničnih kosti v treh smereh (pubococcygeus, iliococcygeus in ishiococcygeus). Ta plast mišic se imenuje tudi medenična diafragma.
Srednja plast mišic se nahaja med simfizo, sramno in sednično kostjo. Srednja plast mišic - urogenitalna diafragma - zavzema sprednjo polovico medeničnega izhoda, skozi katerega potekata sečnica in vagina. V sprednjem delu med njenimi listi so mišični snopi, ki tvorijo zunanji sfinkter sečnice, v zadnjem delu pa so mišični snopi, ki gredo v prečni smeri - globoka prečna mišica presredka.
Spodnja (zunanja) plast mišic medeničnega dna je sestavljena iz površinskih mišic, katerih oblika spominja na številko 8. Sem spadajo bulbokavernozna, ishiokavernozna, zunanja analna mišica zapiralka in površinska transverzalna perinealna mišica.
Ontogeneza jajčnikov
Proces rasti in atrezije foliklov se začne v 20. tednu nosečnosti, do poroda pa v jajčnikih deklice ostane do 2 milijona jajčnih celic. Do menarhe se njihovo število zmanjša na 300 tisoč. V celotnem obdobju reproduktivnega življenja dozori in ovulira največ 500 foliklov. Začetna rast foliklov ni odvisna od stimulacije FSH, je omejena in hitro pride do atrezije. Domneva se, da so namesto steroidnih hormonov glavni regulator rasti in atrezije primarnih foliklov lokalni avtokrini/parakrini peptidi. Domneva se, da procesa rasti in atrezije foliklov ne prekinjajo nobeni fiziološki procesi. Ta proces se nadaljuje v vseh starostnih obdobjih, vključno z intrauterinim obdobjem in menopavzo, prekinejo pa ga nosečnost, ovulacija in anovulacija. Mehanizem, ki sproži rast foliklov in njihovo število v vsakem posameznem ciklu, še vedno ni jasen.
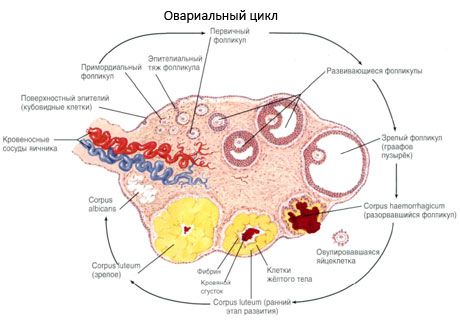
Med svojim razvojem folikel prehaja skozi več razvojnih stopenj. Primordialne zarodne celice izvirajo iz endoderma rumenjakove vrečke, alantoisa, in se v 5.–6. tednu nosečnosti preselijo v genitalno področje zarodka. Zaradi hitre mitotične delitve, ki traja od 6.–8. tedna do 16.–20. tedna nosečnosti, se v jajčnikih zarodka oblikuje do 6–7 milijonov oocitov, obdanih s tanko plastjo granuloznih celic.
Preantralni folikel - oocit je obdan z membrano (Zona pellucida). Granulozne celice, ki obdajajo oocit, se začnejo razmnoževati, njihova rast je odvisna od gonadotropinov in korelira z ravnjo estrogenov. Granulozne celice so tarča za FSH. V fazi preantralnega folikla so granulozne celice sposobne sintetizirati tri razrede steroidov: pretežno inducira aktivnost aromataze, glavnega encima, ki pretvarja androgene v estradiol. Domneva se, da je estradiol sposoben povečati število lastnih receptorjev in tako imeti neposreden mitogeni učinek na granulozne celice neodvisno od FSH. Velja za parakrini dejavnik, ki krepi učinke FSH, vključno z aktivacijo procesov aromatizacije.
Receptorji FSH se pojavijo na membranah granuloznih celic takoj, ko se začne rast folikla. Zmanjšanje ali povečanje FSH vodi do spremembe števila njegovih receptorjev. To delovanje FSH modulirajo rastni faktorji. FSH deluje prek G-proteina, sistema adenilat ciklaze, čeprav steroidogenezo v foliklu v glavnem uravnava FSH, je v ta proces vključenih veliko dejavnikov: ionski kanali, receptorji tirozin kinaze, fosfolipazni sistem sekundarnih prenašalcev.
Vloga androgenov v zgodnjem razvoju foliklov je kompleksna. Granulozne celice imajo androgene receptorje. Ti niso le substrat za aromatizacijo v estrogene, ki jo povzroča FSH, temveč lahko aromatizacijo tudi okrepijo pri nizkih koncentracijah. Ko se raven androgenov poveča, preantralne granulozne celice prednostno ne izberejo poti aromatizacije v estrogene, temveč enostavnejšo androgeno pot preko 5α-reduktaze za pretvorbo v androgen, ki se ne more pretvoriti v estrogen, s čimer zavirajo aktivnost aromataze. Ta proces zavira tudi nastajanje receptorjev za FSH in LH, s čimer ustavi razvoj foliklov.
Pri procesu aromatizacije folikel z visokimi ravnmi androgenov podvrže procesom atrezije. Rast in razvoj folikla sta odvisna od njegove sposobnosti pretvorbe androgenov v estrogene.
V prisotnosti FSH bodo prevladujoča snov folikularne tekočine estrogeni. V odsotnosti FSH - androgeni. LH je običajno odsoten v folikularni tekočini do sredine cikla. Takoj ko se raven LH poveča, se mitotična aktivnost granuloznih celic zmanjša, pojavijo se degenerativne spremembe in raven androgenov v foliklu se poveča. Raven steroidov v folikularni tekočini je višja kot v plazmi in odraža funkcionalno aktivnost jajčnih celic: granuloznih in teka celic. Če je edina tarča za FSH granulozne celice, potem ima LH veliko tarč - teka celice, stromalne in luteinske celice ter granulozne celice. Tako granulozne kot teka celice imajo sposobnost steroidogeneze, vendar v granuloznih celicah prevladuje aromatazna aktivnost.
Kot odgovor na LH tekalne celice proizvajajo androgene, ki jih nato granulozne celice pretvorijo v estrogene z aromatizacijo, ki jo inducira FSH.
Ko folikel raste, tekalne celice začnejo izražati gene za LH receptorje, P450 sec in 3beta-hidroksisteroid dehidrogenazo, inzulinu podoben rastni faktor (IGF-1) sinergistično z LH poveča izražanje genov, vendar ne spodbuja steroidogeneze.
Steroidogeneza v jajčnikih je vedno odvisna od LH. Ko folikel raste, teka celice izražajo encim P450c17, ki iz holesterola tvori androgen. Granulozne celice tega encima nimajo in so za proizvodnjo estrogenov iz androgenov odvisne od teka celic. Za razliko od steroidogeneze je folikulogeneza odvisna od FSH. Ko folikel raste in se raven estrogena povečuje, se aktivira mehanizem povratne zanke – proizvodnja FSH je zavirana, kar posledično vodi do zmanjšanja aktivnosti aromataze v foliklu in na koncu do folikularne atrezije zaradi apoptoze (programirane celične smrti).
Mehanizem povratne zanke estrogenov in FSH zavira razvoj foliklov, ki so začeli rasti, ne pa dominantnega folikla. Dominantni folikel vsebuje več receptorjev FSH, ki podpirajo proliferacijo granuloznih celic in aromatizacijo androgenov v estrogene. Poleg tega parakrina in avtokrina pot delujeta kot pomemben koordinator razvoja antralnih foliklov.
Avtokrini/parakrini regulator sestavljajo peptidi (inhibin, aktivin, folistatin), ki jih sintetizirajo granulozne celice kot odziv na FSH in vstopijo v folikularno tekočino. Inhibin zmanjša izločanje FSH; aktivin spodbuja sproščanje FSH iz hipofize in krepi delovanje FSH v jajčniku; folistatin zavira aktivnost FSH, verjetno z vezavo aktivina. Po ovulaciji in razvoju rumenega telesa je inhibin pod nadzorom LH.
Na rast in diferenciacijo celic jajčnikov vplivajo inzulinu podobni rastni faktorji (IGE). IGF-1 deluje na granulozne celice, kar povzroča povečanje cikličnega adenozin monofosfata (cAMP), progesterona, oksitocina, proteoglikana in inhibina.
IGF-1 deluje na teka celice in povzroča povečano proizvodnjo androgenov. Teka celice nato proizvajajo faktor tumorske nekroze (TNF) in epidermalni rastni faktor (EGF), ki ju prav tako uravnava FSH.
EGF spodbuja proliferacijo granuloznih celic. IGF-2 je glavni rastni faktor v folikularni tekočini, v njej pa so tudi IGF-1, TNF-a, TNF-3 in EGF.
Zdi se, da motnje parakrine in/ali avtokrine regulacije delovanja jajčnikov igrajo vlogo pri motnjah ovulacije in razvoju policističnih jajčnikov.
Z rastjo antralnega folikla se vsebnost estrogena v folikularni tekočini povečuje. Na vrhuncu njihovega povečanja se na granuloznih celicah pojavijo receptorji za LH, pride do luteinizacije granuloznih celic in poveča se proizvodnja progesterona. Tako v predovulatornem obdobju povečana proizvodnja estrogena povzroči pojav LH receptorjev, LH pa nato povzroči luteinizacijo granuloznih celic in proizvodnjo progesterona. Povečanje progesterona zmanjša raven estrogena, kar očitno povzroči drugi vrh FSH sredi cikla.
Ovulacija naj bi se pojavila 10–12 ur po vrhuncu LH in 24–36 ur po vrhuncu estradiola. LH naj bi spodbujal redukcijsko delitev oocitov, luteinizacijo granuloznih celic ter sintezo progesterona in prostaglandina v foliklu.
Progesteron poveča aktivnost proteolitičnih encimov, ki skupaj s prostaglandinom sodelujejo pri razpadu stene folikla. S progesteronom povzročen vrh FSH omogoča oocitu, da izstopi iz folikla s pretvorbo plazminogena v proteolitični encim plazmin, in zagotavlja zadostno število LH receptorjev za normalen razvoj lutealne faze.
V 3 dneh po ovulaciji se granulozne celice povečajo, v njih pa se pojavijo značilne vakuole, napolnjene s pigmentom luteinom. Tekalultealne celice se diferencirajo iz teke in strome ter postanejo del rumenega telesa. Kapilare, ki prodirajo v rumeno telo, se pod vplivom dejavnikov angiogeneze zelo hitro razvijejo, z izboljšano vaskularizacijo pa se poveča proizvodnja progesterona in estrogenov. Aktivnost steroidogeneze in življenjska doba rumenega telesa sta določeni z ravnijo LH. Rumeno telo ni homogena celična tvorba. Poleg dveh vrst lutealnih celic vsebuje endotelijske celice, makrofage, fibroblaste itd. Velike lutealne celice proizvajajo peptide (relaksin, oksitocin) in so bolj aktivne pri steroidogenezi z večjo aktivnostjo aromataze in večjo sintezo progesterona kot majhne celice.
Vrh progesterona se pojavi 8. dan po vrhu LH. Ugotovljeno je, da se progesteron in estradiol v lutealni fazi izločata epizodično, kar je povezano s pulzirajočim izločanjem LH. Z nastankom rumenega telesa se nadzor nad proizvodnjo inhibina prenese s FSH na LH. Inhibin se poveča z zvišanjem estradiola pred vrhom LH in se še naprej povečuje po vrhu LH, čeprav se raven estrogena zmanjša. Čeprav inhibin in estradiol izločajo granulozne celice, ju uravnavajo različne poti. Zmanjšanje inhibina na koncu lutealne faze prispeva k povečanju FSH v naslednjem ciklu.
Rumeno telo se zelo hitro zmanjša - 9.-11. dan po ovulaciji.
Mehanizem degeneracije ni jasen in ni povezan z luteolitično vlogo estrogenov ali z mehanizmom, povezanim z receptorji, kot je opažen v endometriju. Obstaja še ena razlaga za vlogo estrogenov, ki jih proizvaja rumeno telo (corpus luteum). Znano je, da so estrogeni potrebni za sintezo progesteronskih receptorjev v endometriju. Estrogeni lutealne faze so verjetno potrebni za spremembe endometrija, povezane s progesteronom, po ovulaciji. Nezadosten razvoj progesteronskih receptorjev kot posledica nezadostnih ravni estrogena je verjetno dodaten mehanizem za neplodnost in zgodnje izgube nosečnosti, kar je še ena oblika pomanjkanja lutealne faze. Domneva se, da se življenjska doba rumenega telesa določi v času ovulacije. In zagotovo se bo zmanjšala, če je zaradi nosečnosti ne podpira humani horionski gonadotropin. Tako regresija rumenega telesa povzroči zmanjšanje ravni estradiola, progesterona in inhibina. Zmanjšanje inhibina odpravi njegov zaviralni učinek na FSH; Zmanjšanje estradiola in progesterona omogoča zelo hitro obnovo izločanja GnRH in odstranitev mehanizma povratne zanke iz hipofize. Zmanjšanje inhibina in estradiola skupaj s povečanjem GnRH povzroči povečanje FSH namesto LH. Povečanje FSH vodi do rasti foliklov s poznejšo izbiro dominantnega folikla in začne se nov cikel, če ne pride do nosečnosti. Steroidni hormoni igrajo vodilno vlogo v reproduktivni biologiji in splošni fiziologiji. Določajo človeški fenotip, vplivajo na srčno-žilni sistem, presnovo kosti, kožo, splošno počutje in igrajo ključno vlogo pri nosečnosti. Delovanje steroidnih hormonov odraža znotrajcelične in genetske mehanizme, ki so potrebni za prenos zunajceličnega signala v celično jedro, da povzroči fiziološki odziv.
Estrogeni difuzno prodrejo skozi celično membrano in se vežejo na receptorje, ki se nahajajo v celičnem jedru. Kompleks receptor-steroid se nato veže na DNK. V ciljnih celicah te interakcije vodijo do izražanja genov, sinteze beljakovin ter specifičnega delovanja celic in tkiv.