Medicinski strokovnjak članka

Nove publikacije
hipofiza
Zadnji pregled: 04.07.2025

Vsa vsebina iLive je pregledana ali preverjena, da se zagotovi čim večja dejanska natančnost.
Imamo stroge smernice za pridobivanje virov in samo povezave do uglednih medijskih strani, akademskih raziskovalnih institucij in, kadar je to mogoče, medicinsko pregledanih študij. Upoštevajte, da so številke v oklepajih ([1], [2] itd.) Povezave, ki jih je mogoče klikniti na te študije.
Če menite, da je katera koli naša vsebina netočna, zastarela ali drugače vprašljiva, jo izberite in pritisnite Ctrl + Enter.
Hipofiza (hypophysis, s.glandula pituitaria) se nahaja v hipofizni votlini turškega sedla sfenoidne kosti in je od lobanjske votline ločena z odrastkom dura mater možganov, ki tvori diafragmo selle. Skozi odprtino v tej diafragmi je hipofiza povezana z infundibulumom hipotalamusa diencefalona. Prečna velikost hipofize je 10-17 mm, anteroposteriorna 5-15 mm, navpična 5-10 mm. Masa hipofize pri moških je približno 0,5 g, pri ženskah pa 0,6 g. Hipofiza je od zunaj prekrita s kapsulo.
V skladu z razvojem hipofize iz dveh različnih začetkov se v organu razlikujeta dva režnja - sprednji in zadnji. Adenohipofiza ali sprednji reženj (adenohypophysis, s.lobus anterior) je večji in predstavlja 70-80 % celotne mase hipofize. Je gostejši od zadnjega režnja. V sprednjem režnju se razlikuje distalni del (pars distalis), ki zavzema sprednji del hipofizne jame, vmesni del (pars intermedia), ki se nahaja na meji z zadnjim režnjem, in gomoljasti del (pars tuberalis), ki se dviga navzgor in povezuje z infundibulumom hipotalamusa. Zaradi obilice krvnih žil ima sprednji reženj bledo rumeno barvo z rdečkastim odtenkom. Parenhim sprednje hipofize predstavlja več vrst žleznih celic, med katerimi se nahajajo sinusoidne krvne kapilare. Polovica (50 %) celic adenohipofize so kromofilni adenociti, ki imajo v citoplazmi drobnozrnate granule, ki se dobro obarvajo s kromovimi solmi. To so acidofilni adenociti (40 % vseh celic adenohipofize) in bazofilni adenociti (10 %). Bazofilni adenociti vključujejo gonadotropne, kortikotropne in tirotropne endokrinocite. Kromofobni adenociti so majhni, imajo veliko jedro in majhno količino citoplazme. Te celice veljajo za predhodnike kromofilnih adenocitov. Preostalih 50 % celic adenohipofize so kromofobni adenociti.
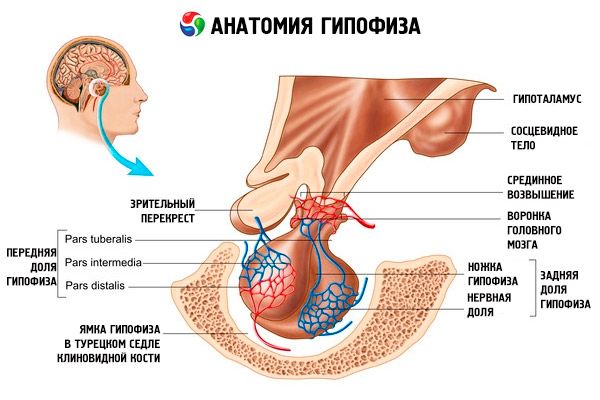
Nevrohipofiza ali zadnji reženj (nevrohipofiza, s.lobus posterior) je sestavljena iz nevronskega režnja (lobus nervosus), ki se nahaja v zadnjem delu hipofizne jame, in lijaka (infundibulum), ki se nahaja za gomoljastim delom adenohipofize. Zadnji reženj hipofize tvorijo nevroglialne celice (celice hipofize), živčna vlakna, ki gredo od nevrosekretornih jeder hipotalamusa do nevrohipofize, in nevrosekretorna telesca.
Hipofiza je preko živčnih vlaken (poti) in krvnih žil funkcionalno povezana s hipotalamusom diencefalona, ki uravnava delovanje hipofize. Hipofiza in hipotalamus skupaj z nevroendokrinimi, žilnimi in živčnimi povezavami običajno veljata za hipotalamo-hipofizni sistem.
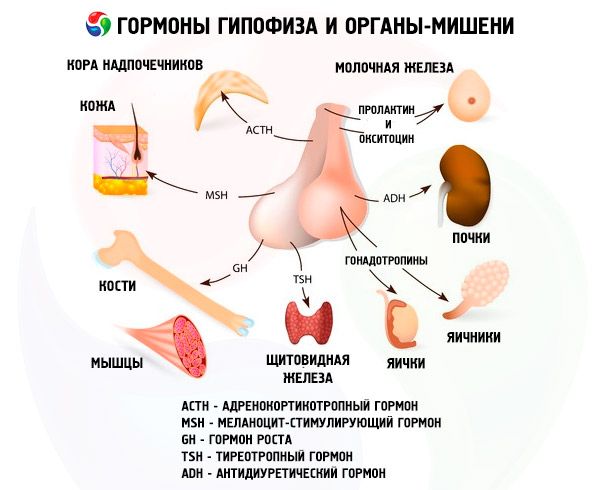
Hormoni sprednje in zadnje hipofize vplivajo na številne funkcije telesa, predvsem prek drugih endokrinih žlez. V sprednji hipofizi acidofilni adenociti (alfa celice) proizvajajo somatotropni hormon (rastni hormon), ki sodeluje pri uravnavanju procesov rasti in razvoja pri mladih organizmih. Kortikotropni endokrinociti izločajo adrenokortikotropni hormon (ACTH), ki spodbuja izločanje steroidnih hormonov nadledvičnih žlez. Tirotropni endokrinociti izločajo tirotropni hormon (TSH), ki vpliva na razvoj ščitnice in aktivira proizvodnjo njenih hormonov. Gonadotropni hormoni: folikle stimulirajoči (FSH), luteinizirajoči (LH) in prolaktin - vplivajo na spolno zorenje telesa, uravnavajo in spodbujajo razvoj foliklov v jajčniku, ovulacijo, rast mlečnih žlez in proizvodnjo mleka pri ženskah, proces spermatogeneze pri moških. Te hormone proizvajajo bazofilni adenociti (beta celice). Tukaj se izločajo tudi lipotropni faktorji hipofize, ki vplivajo na mobilizacijo in izkoriščanje maščob v telesu. V vmesnem delu sprednjega režnja nastaja melanocite stimulirajoči hormon, ki nadzoruje nastajanje pigmentov - melaninov - v telesu.
Nevrosekretorne celice supraoptičnega in paraventrikularnega jedra v hipotalamusu proizvajajo vazopresin in oksitocin. Ta hormona se po aksonih, ki tvorijo hipotalamo-hipofizni trakt, prenašata v celice zadnjega režnja hipofize. Iz zadnjega režnja hipofize te snovi vstopijo v kri. Hormon vazopresin ima vazokonstrikcijski in antidiuretični učinek, zaradi česar se imenuje tudi antidiuretični hormon (ADH). Oksitocin ima stimulativni učinek na kontraktilnost materničnih mišic, povečuje izločanje mleka iz doječe mlečne žleze, zavira razvoj in delovanje rumenega telesa ter vpliva na spremembe tonusa gladkih (neprogastih) mišic prebavil.
Razvoj hipofize
Sprednji reženj hipofize se razvije iz epitelija hrbtne stene ustnega prekata v obliki obročaste izrastke (Rathkejeva vrečka). Ta ektodermalna izboklina raste proti dnu bodočega tretjega prekata. Proti njemu, od spodnje površine drugega možganskega mehurčka (bodoče dno tretjega prekata), raste odrastek, iz katerega se razvijeta sivi tuberkul infundibuluma in zadnji reženj hipofize.
Plovila in živci hipofize
Iz notranjih karotidnih arterij in žil arterijskega kroga velikih možganov se zgornja in spodnja hipofizna arterija usmerita v hipofizo. Zgornje hipofizne arterije gredo v sivo jedro in infundibulum hipotalamusa, se tukaj anastomozirajo in tvorijo kapilare, ki prodirajo v možgansko tkivo - primarno hemokapilarno mrežo. Iz dolgih in kratkih zank te mreže se oblikujejo portalne vene, ki so usmerjene v sprednji reženj hipofize. V parenhimu sprednjega režnja hipofize se te vene razdelijo v široke sinusoidne kapilare in tvorijo sekundarno hemokapilarno mrežo. Zadnji reženj hipofize se s krvjo oskrbuje predvsem iz spodnje hipofizne arterije. Med zgornjo in spodnjo hipofizno arterijo so dolge arterijske anastomoze. Odtok venske krvi iz sekundarne hemokapilarne mreže poteka skozi sistem ven, ki se izlivajo v kavernozne in interkavernozne sinuse dure mater.
Simpatična vlakna, ki vstopajo v organ skupaj z arterijami, sodelujejo pri inervaciji hipofize. Postganglijska simpatična živčna vlakna odstopajo od pleksusa notranje karotidne arterije. Poleg tega se v zadnjem režnju hipofize nahajajo številni konci procesov nevrosekretornih celic, ki se nahajajo v jedrih hipotalamusa.
Starostne značilnosti hipofize
Povprečna teža hipofize pri novorojenčkih doseže 0,12 g. Teža organa se do 10. leta starosti podvoji, do 15. leta pa potroji. Do 20. leta starosti teža hipofize doseže svoj maksimum (530–560 mg) in v naslednjih starostnih obdobjih ostane skoraj nespremenjena. Po 60. letu opazimo rahlo zmanjšanje teže te endokrine žleze.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hipofizni hormoni
Enotnost živčne in hormonske regulacije v telesu zagotavlja tesna anatomska in funkcionalna povezava hipofize in hipotalamusa. Ta kompleks določa stanje in delovanje celotnega endokrinega sistema.
Glavna endokrina žleza, ki proizvaja številne peptidne hormone, ki neposredno uravnavajo delovanje perifernih žlez, je hipofiza. Je rdečkasto-siva tvorba v obliki fižola, prekrita z vlaknasto kapsulo, ki tehta 0,5-0,6 g. Njena velikost se nekoliko razlikuje glede na spol in starost osebe. Splošno sprejeta delitev hipofize na dva režnja, ki se razlikujeta po razvoju, strukturi in funkciji: sprednji distalni - adenohipofiza in zadnji - nevrohipofiza. Prvi predstavlja približno 70 % celotne mase žleze in je običajno razdeljen na distalni, infundibularni in vmesni del, drugi - na zadnji del oziroma reženj in steblo hipofize. Žleza se nahaja v hipofizični jami turškega sedla sfenoidne kosti in je preko stebla povezana z možgani. Zgornji del sprednjega režnja pokriva optična hiazma in optični trakti. Prekrvavitev hipofize je zelo obilna in jo zagotavljajo veje notranje karotidne arterije (zgornja in spodnja hipofizna arterija) ter veje arterijskega kroga velikih možganov. Zgornje hipofizne arterije sodelujejo pri prekrvavitvi adenohipofize, spodnje pa nevrohipofize, pri čemer se stikajo z nevrosekretornimi končiči aksonov velikih celičnih jeder hipotalamusa. Prve vstopijo v medialno eminenco hipotalamusa, kjer se razpršijo v kapilarno mrežo (primarni kapilarni pletež). Te kapilare (s katerimi se stikajo končiči aksonov majhnih nevrosekretornih celic mediobazalnega hipotalamusa) se zbirajo v portalne vene, ki se vzdolž stebla hipofize spuščajo v parenhim adenohipofize, kjer se spet razdelijo v mrežo sinusoidnih kapilar (sekundarni kapilarni pletež). Tako kri, ki je predhodno prešla skozi srednjo eminenco hipotalamusa, kjer se obogati s hipotalamičnimi adenohipofiziotropnimi hormoni (sproščujočimi hormoni), vstopi v adenohipofizo.
Odtok krvi, nasičene z adenohipofiznimi hormoni, iz številnih kapilar sekundarnega pleksusa poteka skozi sistem ven, ki se nato izlivajo v venske sinuse dura mater in nato v splošni krvni obtok. Tako je portalni sistem hipofize s padajočo smerjo pretoka krvi iz hipotalamusa morfofunkcionalna komponenta kompleksnega mehanizma nevrohumoralne regulacije tropskih funkcij adenohipofize.
Hipofizo oživčujejo simpatična vlakna, ki sledijo hipofiznim arterijam. Izvirajo iz postganglijskih vlaken, ki potekajo skozi notranji karotidni pleksus in so povezana z zgornjimi cervikalnimi gangliji. Adenohipofiza nima neposredne oživitve iz hipotalamusa. Zadnji reženj prejema živčna vlakna iz nevrosekretornih jeder hipotalamusa.
Adenohipofiza je v svoji histološki arhitekturi zelo kompleksna tvorba. Ima dve vrsti žleznih celic - kromofobne in kromofilne. Slednje se nato delijo na acidofilne in bazofilne (podroben histološki opis hipofize je podan v ustreznem poglavju priročnika). Vendar je treba opozoriti, da se hormoni, ki jih proizvajajo žlezne celice, ki sestavljajo parenhim adenohipofize, zaradi raznolikosti slednje do neke mere razlikujejo po svoji kemijski naravi, fina struktura izločajočih celic pa mora ustrezati biosintetskim značilnostim vsake od njih. Včasih pa lahko v adenohipofizi opazimo prehodne oblike žleznih celic, ki so sposobne proizvajati več hormonov. Obstajajo dokazi, da vrsta žleznih celic adenohipofize ni vedno genetsko določena.
Pod diafragmo turškega sedla se nahaja lijakasti del sprednjega režnja. Objema pecelj hipofize in se dotika sivega izrastka. Ta del adenohipofize je značilen po prisotnosti epitelijskih celic in obilni prekrvavitvi. Je tudi hormonsko aktiven.
Vmesni (srednji) del hipofize je sestavljen iz več plasti velikih sekretorno aktivnih bazofilnih celic.
Hipofiza opravlja različne funkcije s pomočjo svojih hormonov. Njen sprednji reženj proizvaja adrenokortikotropni (ACTH), tirotropni (TSH), folikle stimulirajoči (FSH), luteinizirajoči (LH), lipotropni hormon, pa tudi rastni hormon - somatotropni (STO) in prolaktin. V vmesnem režnju se sintetizira melanocite stimulirajoči hormon (MSH), v zadnjem režnju pa se kopičita vazopresin in oksitocin.
ACTH
Hipofizni hormoni so skupina beljakovinskih in peptidnih hormonov ter glikoproteinov. Od hormonov sprednje hipofize je ACTH najbolje raziskan. Proizvajajo ga bazofilne celice. Njegova glavna fiziološka funkcija je spodbujanje biosinteze in izločanja steroidnih hormonov s skorjo nadledvične žleze. ACTH kaže tudi melanocitno stimulirajočo in lipotropno aktivnost. Leta 1953 je bil izoliran v čisti obliki. Kasneje je bila ugotovljena njegova kemijska struktura, ki jo sestavlja 39 aminokislinskih ostankov pri ljudeh in številnih sesalcih. ACTH nima vrstne specifičnosti. Trenutno je bila izvedena kemična sinteza tako samega hormona kot različnih fragmentov njegove molekule, ki so bolj aktivni kot naravni hormoni. Struktura hormona ima dva dela peptidne verige, od katerih eden zagotavlja detekcijo in vezavo ACTH na receptor, drugi pa zagotavlja biološki učinek. Očitno se veže na receptor ACTH z interakcijo električnih nabojev hormona in receptorja. Vlogo biološkega efektorja ACTH opravlja fragment molekule 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
Melanocitno stimulirajoča aktivnost ACTH je posledica prisotnosti N-terminalne regije v molekuli, ki jo sestavlja 13 aminokislinskih ostankov in ponavlja strukturo alfa-melanocitno stimulirajočega hormona. Ta regija vsebuje tudi heptapeptid, ki je prisoten v drugih hipofiznih hormonih in ima nekatere adrenokortikotropne, melanocitno stimulirajoče in lipotropne aktivnosti.
Ključni trenutek v delovanju ACTH velja za aktivacijo encima protein kinaze v citoplazmi s sodelovanjem cAMP. Fosforilirana protein kinaza aktivira encim esterazo, ki v maščobnih kapljicah pretvori holesterolske estre v prosto snov. Beljakovina, sintetizirana v citoplazmi kot posledica fosforilacije ribosomov, spodbuja vezavo prostega holesterola na citokrom P-450 in njegov prenos iz lipidnih kapljic v mitohondrije, kjer so prisotni vsi encimi, ki zagotavljajo pretvorbo holesterola v kortikosteroide.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Ščitnično stimulirajoči hormon
TSH - tirotropin - glavni regulator razvoja in delovanja ščitnice, procesov sinteze in izločanja ščitničnih hormonov. Ta kompleksna beljakovina - glikoprotein - je sestavljena iz alfa in beta podenot. Struktura prve podenote sovpada z alfa podenoto luteinizirajočega hormona. Poleg tega se v veliki meri ujema pri različnih živalskih vrstah. Zaporedje aminokislinskih ostankov v beta podenoti človeškega TSH je bilo dešifrirano in je sestavljeno iz 119 aminokislinskih ostankov. Opazimo lahko, da so si beta podenote človeškega in govejega TSH v marsičem podobne. Biološke lastnosti in narava biološke aktivnosti glikoproteinskih hormonov so določene z beta podenoto. Zagotavlja tudi interakcijo hormona z receptorji v različnih ciljnih organih. Vendar pa beta podenota pri večini živali kaže specifično aktivnost šele po združitvi z alfa podenoto, ki deluje kot nekakšen aktivator hormona. Slednja z enako verjetnostjo sproži luteinizirajočo, folikle stimulirajočo in tirotropno aktivnost, ki jo določajo lastnosti beta podenote. Ugotovljena podobnost nam omogoča sklepati, da so ti hormoni nastali v procesu evolucije iz enega skupnega predhodnika, beta podenota pa določa tudi imunološke lastnosti hormonov. Obstaja predpostavka, da alfa podenota ščiti beta podenoto pred delovanjem proteolitičnih encimov in olajša njen transport iz hipofize v periferne ciljne organe.
Gonadotropni hormoni
Gonadotropini so v telesu prisotni v obliki LH in FSH. Funkcionalni namen teh hormonov se običajno zreducira na zagotavljanje reproduktivnih procesov pri posameznikih obeh spolov. Tako kot TSH so kompleksni proteini - glikoproteini. FSH povzroča zorenje foliklov v jajčnikih žensk in spodbuja spermatogenezo pri moških. LH povzroči rupturo foliklov pri ženskah z nastankom rumenega telesca in spodbuja izločanje estrogenov in progesterona. Pri moških isti hormon pospešuje razvoj intersticijskega tkiva in izločanje androgenov. Učinki gonadotropinov so medsebojno odvisni in se pojavljajo sinhrono.
Dinamika izločanja gonadotropinov pri ženskah se med menstrualnim ciklom spreminja in je bila dovolj podrobno preučena. V predovulacijski (folikularni) fazi cikla je vsebnost LH na precej nizki ravni, medtem ko je FSH povišana. Z zorenjem folikla se poveča izločanje estradiola, kar spodbuja povečano proizvodnjo gonadotropinov s strani hipofize in pojav ciklov LH in FSH, tj. spolni steroidi spodbujajo izločanje gonadotropinov.
Trenutno je struktura LH določena. Tako kot TSH je sestavljen iz dveh podenot: a in b. Struktura alfa podenote LH pri različnih živalskih vrstah se v veliki meri ujema, ustreza strukturi alfa podenote TSH.
Struktura beta podenote LH se bistveno razlikuje od strukture beta podenote TSH, čeprav ima štiri enake dele peptidne verige, ki jih sestavljajo 4-5 aminokislinski ostanki. V TSH so lokalizirani na položajih 27-31, 51-54, 65-68 in 78-83. Ker beta podenota LH in TSH določa specifično biološko aktivnost hormonov, lahko domnevamo, da bi morali homologni odseki v strukturi LH in TSH zagotavljati povezavo beta podenot z alfa podenoto, odseki, ki se razlikujejo po strukturi, pa bi morali biti odgovorni za specifičnost biološke aktivnosti hormonov.
Nativni LH je zelo stabilen na delovanje proteolitičnih encimov, vendar beta podenoto hitro cepi kimotripsin, a podenoto pa encim težko hidrolizira, tj. igra zaščitno vlogo, saj preprečuje kimotripsinu dostop do peptidnih vezi.
Kar zadeva kemijsko strukturo FSH, raziskovalci še niso dobili dokončnih rezultatov. Tako kot LH je tudi FSH sestavljen iz dveh podenot, vendar se beta podenota FSH razlikuje od beta podenote LH.
Prolaktin
Drug hormon, prolaktin (laktogeni hormon), igra aktivno vlogo v procesih razmnoževanja. Glavne fiziološke lastnosti prolaktina pri sesalcih se kažejo v obliki spodbujanja razvoja mlečnih žlez in laktacije, rasti lojnic in notranjih organov. Spodbuja manifestacijo učinka steroidov na sekundarne spolne značilnosti pri samcih, spodbuja sekretorno aktivnost rumenega telesa pri miših in podganah ter sodeluje pri uravnavanju presnove maščob. V zadnjih letih se prolaktinu posveča veliko pozornosti kot regulatorju materinskega vedenja; takšno polifunkcionalnost pojasnjuje njegov evolucijski razvoj. Je eden najstarejših hipofiznih hormonov in ga najdemo celo pri dvoživkah. Trenutno je struktura prolaktina pri nekaterih vrstah sesalcev popolnoma dešifrirana. Vendar pa so znanstveniki do nedavnega dvomili o obstoju takega hormona pri ljudeh. Mnogi so verjeli, da njegovo funkcijo opravlja rastni hormon. Zdaj so pridobljeni prepričljivi dokazi o prisotnosti prolaktina pri ljudeh in njegova struktura je delno dešifrirana. Prolaktinski receptorji se aktivno vežejo na rastni hormon in placentni laktogen, kar kaže na en sam mehanizem delovanja treh hormonov.
Somatotropin
Rastni hormon, somatotropin, ima še širši spekter delovanja kot prolaktin. Tako kot prolaktin ga proizvajajo acidofilne celice adenohipofize. STH spodbuja rast skeleta, aktivira biosintezo beljakovin, ima učinek mobilizacije maščob in spodbuja povečanje telesne mase. Poleg tega usklajuje presnovne procese.
Sodelovanje hormona v slednjem potrjuje dejstvo močnega povečanja njegovega izločanja s strani hipofize, na primer, ko se raven sladkorja v krvi zniža.
Kemijska struktura tega človeškega hormona je zdaj v celoti ugotovljena – 191 aminokislinskih ostankov. Njegova primarna struktura je podobna strukturi horionskega somatomamotropina ali placentnega laktogena. Ti podatki kažejo na pomembno evolucijsko bližino obeh hormonov, čeprav kažeta razlike v biološki aktivnosti.
Poudariti je treba visoko vrstno specifičnost obravnavanega hormona – na primer, STH živalskega izvora je pri ljudeh neaktiven. To je razloženo tako z reakcijo med receptorji STH pri ljudeh in živalih kot tudi s samo strukturo hormona. Trenutno potekajo študije za identifikacijo aktivnih centrov v kompleksni strukturi STH, ki kažejo biološko aktivnost. Preučujejo se posamezni fragmenti molekule, ki kažejo druge lastnosti. Na primer, po hidrolizi človeškega STH s pepsinom je bil izoliran peptid, ki je sestavljen iz 14 aminokislinskih ostankov in ustreza odseku molekule 31–44. Ni imel rastnega učinka, je pa znatno presegel naravni hormon v lipotropni aktivnosti. Človeški rastni hormon ima za razliko od podobnega živalskega hormona pomembno laktogeno aktivnost.
Adenohipofiza sintetizira številne peptidne in beljakovinske snovi, ki imajo učinek mobilizacije maščob, lipotropni učinek pa imajo hipofizni tropni hormoni - ACTH, STH, TSH in drugi. V zadnjih letih so še posebej poudarjeni beta- in γ-lipotropni hormoni (LPG). Biološke lastnosti beta-LPG so bile najbolj podrobno preučene; poleg lipotropnega delovanja ima tudi melanocitno stimulirajoči, kortikotropin stimulirajoči in hipokalcemični učinek, povzroča pa tudi inzulinu podoben učinek.
Trenutno je dešifrirana primarna struktura ovčjega LPG (90 aminokislinskih ostankov), lipotropnih hormonov prašičev in goveda. Ta hormon ima vrstno specifičnost, čeprav je struktura osrednje regije beta-LPG pri različnih vrstah enaka. To določa biološke lastnosti hormona. Eden od fragmentov te regije se nahaja v strukturi alfa-MSH, beta-MSH, ACTH in beta-LPG. Domneva se, da so ti hormoni v procesu evolucije nastali iz istega predhodnika. γ-LPG ima šibkejšo lipotropno aktivnost kot beta-LPG.
Melanocitno stimulirajoči hormon
Ta hormon, sintetiziran v vmesnem režnju hipofize, spodbuja biosintezo kožnega pigmenta melanina pri njegovi biološki funkciji, spodbuja povečanje velikosti in števila pigmentnih celic melanocitov v koži dvoživk. Te lastnosti MSH se uporabljajo pri biološkem testiranju hormona. Obstajata dve vrsti hormona: alfa- in beta-MSH. Dokazano je, da alfa-MSH nima vrstne specifičnosti in ima enako kemijsko strukturo pri vseh sesalcih. Njegova molekula je peptidna veriga, sestavljena iz 13 aminokislinskih ostankov. Beta-MSH pa ima vrstno specifičnost in se njegova struktura razlikuje pri različnih živalih. Pri večini sesalcev molekula beta-MSH sestoji iz 18 aminokislinskih ostankov, le pri ljudeh pa je od aminokislinskega konca podaljšana za štiri aminokislinske ostanke. Treba je opozoriti, da ima alfa-MSH nekaj adrenokortikotropnega delovanja, njegov vpliv na vedenje živali in ljudi pa je zdaj dokazan.
Oksitocin in vazopresin
Vasopresin in oksitocin, ki se sintetizirata v hipotalamusu, se kopičita v zadnjem režnju hipofize: vazopresin v nevronih supraoptičnega jedra, oksitocin pa v paraventrikulacijskem jedru. Nato se preneseta v hipofizo. Poudariti je treba, da se predhodnik hormona vazopresina najprej sintetizira v hipotalamusu. Hkrati se tam proizvajata tudi proteina nevrofizin tipa 1 in 2. Prvi se veže na oksitocin, drugi pa na vazopresin. Ta kompleksa se v obliki nevrosekretornih granul selita v citoplazmi vzdolž aksona in dosežeta zadnji reženj hipofize, kjer se živčna vlakna končajo v žilni steni in vsebina granul vstopi v kri. Vasopresin in oksitocin sta prva hipofizna hormona s popolnoma uveljavljenim zaporedjem aminokislin. Po svoji kemijski strukturi sta nonapeptida z enim disulfidnim mostom.
Obravnavani hormoni imajo različne biološke učinke: spodbujajo transport vode in soli skozi membrane, imajo vazopresorski učinek, krepijo krčenje gladkih mišic maternice med porodom in povečujejo izločanje mlečnih žlez. Treba je opozoriti, da ima vazopresin večjo antidiuretično aktivnost kot oksitocin, medtem ko ima slednji močnejši učinek na maternico in mlečno žlezo. Glavni regulator izločanja vazopresina je poraba vode; v ledvičnih tubulih se veže na receptorje v citoplazemskih membranah, čemur sledi aktivacija encima adenilat ciklaze v njih. Za vezavo hormona na receptor in za biološki učinek so odgovorni različni deli molekule.
Hipofiza, ki je preko hipotalamusa povezana s celotnim živčnim sistemom, združuje endokrini sistem v funkcionalno celoto in sodeluje pri zagotavljanju stalnosti notranjega okolja telesa (homeostaza). Znotraj endokrinega sistema se homeostatska regulacija izvaja na podlagi načela povratne zveze med sprednjim režnjem hipofize in "ciljnimi" žlezami (ščitnica, nadledvična skorja, spolne žleze). Presežek hormona, ki ga proizvaja "ciljna" žleza, zavira, njegovo pomanjkanje pa spodbuja izločanje in sproščanje ustreznega tropnega hormona. Hipotalamus je vključen v sistem povratne zveze. V njem se nahajajo receptorske cone, občutljive na hormone "ciljnih" žlez. S specifično vezavo na hormone, ki krožijo v krvi, in spreminjanjem odziva glede na koncentracijo hormonov hipotalamični receptorji prenašajo svoj učinek na ustrezne hipotalamične centre, ki usklajujejo delo adenohipofize in sproščajo hipotalamične adenohipofiziotropne hormone. Zato je treba hipotalamus obravnavati kot nevroendokrini možgani.